|
|
|
|
|
|
|
|
|
|
 | Diversity of vascular plants in Central-Europe (CE |
|
|
Contents of this page
|
|
|
|
Top_________________________Next
|
Introduction
|
 |
The central part of Europe would be covered almost entirely by a summer-green deciduous beech and oak forest. However, Central-Europe is an ancient cultivated landscape and the present-day vegetation as a result of millennia of mankind's history has literally nowhere retained its original chareacter (Ellenberg & Strutt 2009).
|
|
|
Nowadays, the Central-European countriyside consists of a mosaic of planted forests, pastures, meadows and arable land. Urbanization and mass transportation as a consequence of the dense humam population as well as industrialization contirubute further to the fragmentation of the landscape (Ellenberg & Strutt 2009).
|
|
|
In this section the German Red List serves as a proxy for the threat status of vascular plants in Central-Europe. Detailied information contained in the Red List allows for an analysis of causes underlying the status in major vegetation fomrations.
|
|
|
|
|
|
|
Top_________________________Next
|
Threat status of vasular plants
|
|
|
According to the Red List the German Flora contains 3001 native species of vascular plants. Another 318 are neophytes, which have entered the region after 1492 and are ignored in the calculations, though. Figure 2 displays that 943 species of vascular plants are currently either extinct or threatened with extinction in Germany (compare Threat situation of plant groups in Germany). With a fraction of 31.4% threatened species the situation is substantially inferiior to that of the global scale.
|
|
|
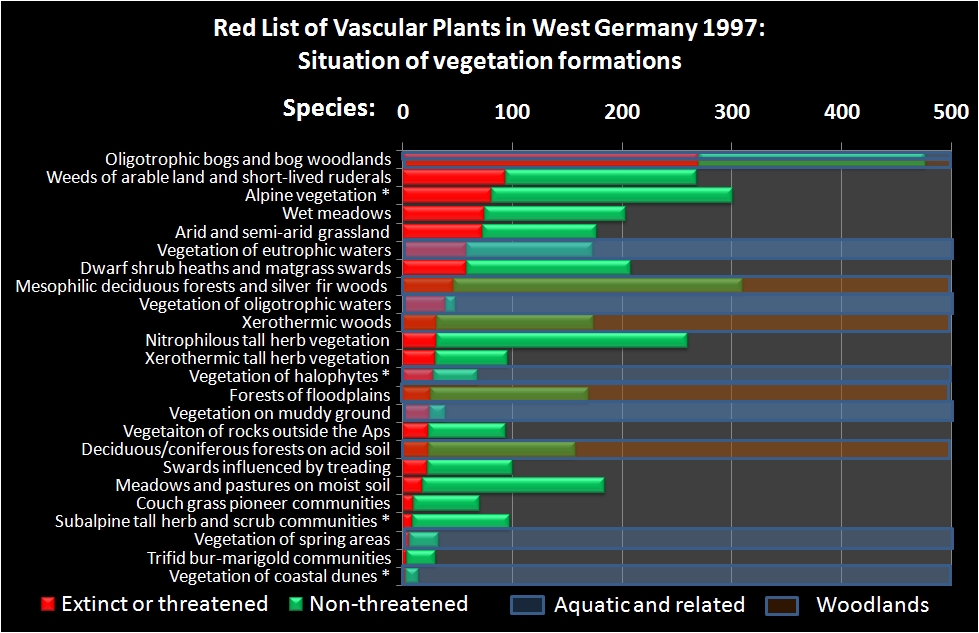
A closer look at the status in the 24 German vegetation formations reveals that "Oligotrophic bogs and bog woodlands" is the formation with the highest species richness. Besides that, it contains by far the highest number of extinct or threatened species (see Figure 3, compare Table 6 in Jedicke 1997, Slide 4 in Marx et al. 1997).
|
|
|
The next four formations following in the ranking are: "Weed communities of arable land and short-lived ruderal communities", "Alpine vegetation", "Wet medows" and "Arid and semi-arid grasslands". Thus, with the exception of the "Oligotrrophic bogs and bog woodlands" the other top five positions are all held by terrestrial herbaceous communities.
|
|
|
Formations under forest management altogether contain about 130 extinct or thretened species - a rather low number when compared with the roughly 650 species covered by pure terrestrial herbaceous communities. Among the herbaceous communities grasslands with about 380 species hold a major fraction of the vascular plant diversity.
|
|
|
|
|
|
|
Top_________________________Next
|
Causes of the loss in vascular plant diversity
|
|
|
In essence, Table 2 resembles the figure of global biodiversity loss: Land use change is the most important direct driver, followed by nutrient load & pollution and climate change - whose influence is dealt with in the next paragraph.
|
|
|
Under current agro-economic conditions abandonment of extensively-managed area is a widespread factor of land use change in Central-Europe. Usually, those areas are at risk to fall victim to succession afterwards (compare Others colum in Table 2 and Figure 4).
|
|
|
The other prominent factor of land use change is intensification.
|
|
|
Often land use change is understood as a factor that works in isolation. However, here the tight association between land use change and nutrient load is emphasized. The change to more intense management certainly requires larger amounts of nitrogen fertilizer to be applied. Hence, in the three formations where intensifcation was referrred to in the first place as having a strong impact, such an impact was also inferred to nutrient load in Table 2.
|
|
|
In wet formations intensification of management is often accompanied by rainage.
|
|
|
Further, in virtually all formations deposition of air-borne nitrogen contributes, although to a lesser extent, to the nutruent load. At the wet end of the range spanned by the formations lateral N-input via the soil water pathway also plays a significant role.
|
|
|
Obviously, in several vegetation formations, incluing "Alpine vegetation", various leisure activities and tourism exert a damaging influence.
|
|
|
Noteworthy, unlike the global situation economic growth is the only root cause underlying all the direct drivers of diversity loss in Central-Europe, since population growth is absent here.
|
|
|
Table 2 has been adopted from Riecken, Ries & Ssymank (1994) after allocating vegetation formations to respective biotop types.
|
|
Table 2: Direct drivers of threatening
|
|
|
Land use change:
|
Nutrient loading & pollution:
|
Climate change:
|
Other:
|
|
2
|
- - / - - / intensification, abandonment
|
- - / - - / nitrogen fertilizer, herbicides
|
- / - -
|
succession
|
|
3
|
|
- / - / nitrogen deposition from atmosphere
|
- (-) / - (-)
|
intense tourism, skiing
|
|
4
|
- - / - - / abandonment, intensifiction
|
- / - / nitrogen fertilizer, nitrogen input from adjacent sites, nitrogen deposition
|
- (-) / - (-)
|
drainage, lowering of groundwater, river training, succession
|
|
5
|
- - / - - / abandonment, intensification, afforestation
|
- / - / nitrogen fertilizer, nitrogen deposition
|
- (-) / - (-)
|
succession, tourism, trampling, plant collection
|
|
7
|
- - / - - / intesification, abandonment
|
- - / - - / nitrogen fertilizer, nitrogen deposition
|
- (-) / - (-)
|
accelerated succession, intense tourism
|
|
11
|
- / - / intensifiction, afforestation
|
- / - / nitrogen fertilizer, nitrogen deposition
|
- (-) / - (-)
|
river training, loweing of groundwater, drainage
|
|
12
|
|
-/-/ nitrogen deposition
|
- (-) / - (-)
|
stone quarry, climbing sport
|
|
16
|
|
- / - / nitrogen deposition
|
- (-) / - (-)
|
stone quarry, climbing sport
|
|
19
|
- - / - - / intensification, conversion to arable land, conversion of meadows to pasture, afforestation, abandonment
|
- - / - - / nitrogen fertilizer, nitrogen deopsition
|
- (-) / - (-)
|
drainage, house construction, succession
|
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
|
|
|
|
Top_________________________Next
|
Diversity of vascular plants and climate change
|
|
|
Climate change is the third important direct rdriver to loss in CE vascular plant diversity.
|
|
|
The Millenium Ecosystem Assessment (MA 2005) came to the conclusion that biodiversity of the alpine biome is already suffering severely from climate change, wehereas temperate biomes are currently affected only moderately. However, there is widespread consensus that the impact of climate change will rise in the decades to come. Recent work of Thomas et al. (2004) even suggested that climate will become a threat as serious as land use cahnge - but of course this estimate is based on model scenarios with uncertainty.
|
|
|
Meanwhile, the European Environment Agency reports observed climate change impacts on European biota at regular intervals (EEA 2004, EEA 2008).
|
|
|
Among them are phenological changes, such as earlier budbreak and flowering. Phenological alterations are usually measured on a set of common species using a standard protocol.
|
|
|
In contrast, other studies cited by EEA are more compelling, at leat to me, since they are based on natural vegetation under the influence of the whole set of natural environmental factors. They commonly report abundance shifts either at the margin or at the core of species' ranges. Upward and nothward range shifts have been reported frequently, although the ladder results were considered to be inconclusive. Within long-term monitoring programs plant species with Ellenberg indicator values for high temperature became more abundant in Norway and The Netherlands, thereby including CE.
|
|
|
While all these studies give indirect evidence of climate change effects, they do not, however, contribute to our understanding of how and to what extent various climate change factors, such as sunlight, temperature, atmospheric carbon dioxide level, humidity and precipitation, affect the abundance of a species or, more generally speaking, the composition of a plant community.
|
|
|
For instance, subtle CO2-induced changes in the community struccture of moist meadows are likely to occur in Central-Europe according to results from the Giessen Free Air CO2 Enrichment (FACE) experiment (Grueters et al. 2006).
|
|
|
|
Top_________________________Next
|
Conclusion
|
|
|
|
|
|
The major portion of the terrestrial plant diversity in Central-Europe is located in herbaceous communities, particularly in grassland communities. Since UIBM aims to contribute to our understanding of vascular plant diversity it shall simulate the vegetation dynamics within grassland communities, though.
|
|
|
As the graphical summary of Figure 4 suggests UIBM will cover the main causal chain leading to loss in plant diversity by simulating how nutrient load interacts with management intensification and major climate change factors.
|
|
Navigator
|
You are here: Cental-European Plants -> Background -> Home
Local Navigator: Λ Top <- Previous -> Next
Sub-Page Navigator: Biodiversity <- Central-Eurpean Plants -> Modeling Biodiversity -> Future Biodiversity Models -> Online Databases -> Universal Scaling Laws
Whole-Page Navigator: Home <- Background -> UIBM Development -> Virtual Experiments -> Outlook: Virtual Ecology
|
Copyright © Nov. 2009 Dr. U. Grueters
|
|
|
Printable Version
|
|
|
|
|
|
|
|