|
|
|
 | The Universal Individual-Based Model (UIBM) |
|
|
|
|
|
|
|
|
|
Click here to watch UIBM-screenshots enlarged
|
Overview
|
|
UIBM - The Next Generation of Biodiversity Models
|
|
Basic UIBM Idea
|
(which makes UIBM a biodiversity model)
Once a template species is successfully constructed from databases and a Universal Individual-Based Model is developed along that way, more species can be easily constructed, since information in the databases is idential for all species.
|
UIBM Characteristics
|
 |
UIBM aims to simulate plant diversity in herbaceous, grassland communities, which hold the majority of vascular plant diversity in Central-Europe.
|
|
|
UIBM aims to simulate the plant diversity response to management intensification, nutrient load and climate change factors. It improves species distribution models, which restrict themselves to climate change.
|
|
|
UIBM is a process-based model. This is an advantage over empirical/statistical species distribution models, which rely heavily on expert knowledge and suffer from methodological limitations.
|
|
|
UIBM simulates local species composition of herb communities on a Braun-Blanquet minimum area. These are advantages over process-based dynamic global vegetation models, which are limited to large-scale plant functional type responses of natural vegetation and difficult to validate.
|
|
|
UIBM is an individual-/agent-based model with a functional-structural basis.
|
|
|
As with dynamic global vegetation models the functional basis consists of widespread organ energy-/gas-exchange models, scaled up to the canopy via a layered radiation interception and turbulent transfer model.
|
|
|
In UIBM species are constructed from life-cycle traits contained in databases. This better satisfies the huge data requirements of individual-based models on the individual level.
|
|
|
The structural basis is derived from trait minima/maxima on the organ level This replaces data requiremtns of individual-based models on the individual level.
|
|
|
The methodology to construct species in UIBM relies on multivariate allometry and serial biological reasoinng.
|
|
UIBM Parametrization
|
|
|
(experience with Arrhenatherum elatius)
|
|
|
16 plant traits were successfully derived from databases.
|
|
|
9 plant traits could not be derived from databases. Important traits that were not available from the databases were the number of internodes per stem and the root:shoot ratio. Among them were also 5 functional ecophysiological traits. However, I assume that these model parameters could be derived from the literature for many other species.
|
|
|
Moreover, 15 assumptions about plant traits had to be made..
|
|
UIBM Application
|
|
|
Generate locally validated virtual communities in UIBM.
|
|
|
Remove dependence on expert knowledge from species distribution models development. Develop species distribution models from „transparent“ virtual communities generated with UIBM rather than from „black-box“ real communities.
|
|
|
Do virtual climate scenario and management scenario experiments in UIBM.
|
|
|
Do bio-manipulative experiments in UIBM to study species interactions.
|
|
UIBM-Flyer
|
|
Demo-Video 2
|
Click on the figure to watch the template species Arrhenatherum elatius grow in the virtual world. In the demo-video the ontogeny is followed from seedling to the time of first flowering.
|
|
|
Demo-Video 3
|
Click on the figure to watch a virtual vegetation survey plot with 1,000 Arrhenatherum elatius plants.
The 1,000 plants are contained in 100 databases. At an interval of milliseconds of simulatin time 10 plants are queried from a database, stepped and then committed again to the same database. At the end of the whole sequence shown in this video simulation time proceeds to the next time step (in this case: + 1 hr).
|
|
|
UIBM-Screenshots: Enlarge When Clicked
|
|
Starting UIBM
|
|
Doing Virtual Experiments in UIBM
|
|
Inspecting Functions in UIBM
|
|
Inspecting Component Structure in UIBM
|
|
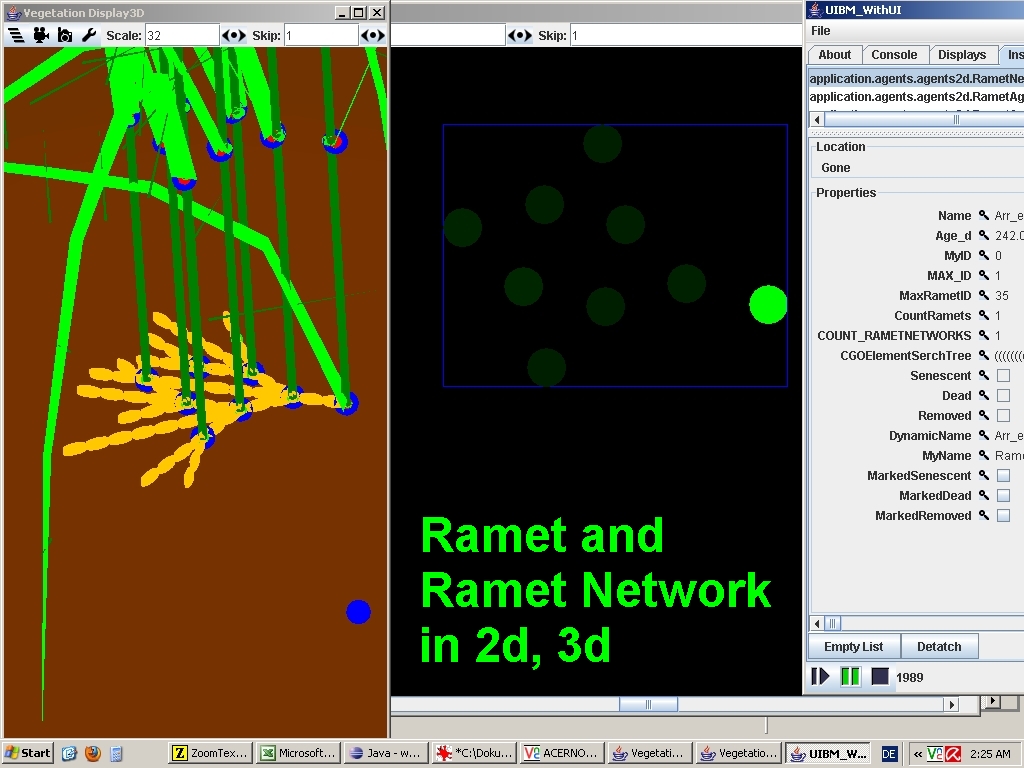
Inspecting Ramets & RametNetworks
|
|
Project Administration & Model Parameterization
|
Dr. U. Grueters
|
Model Design/Implementation
|
Uwe Grueters, uwegrueters@users.sourceforge.net
Roland Dahlem, roland.dahlem@mni.fh-giessen.de
Jochen Senkbeil, jochen.k.senkbeil@bot2.bio.uni-giessen.de
Markus Woetzel, markus@woetzel.net
|
CREDITS & REFERENCES
|
To refer to this model in academic publications, please use:
Uwe Grueters, Roland Dahlem, Jochen Senkbeil, Markus Woetzel (2007):
The Universal Individual-Based Model (UIBM)
http://sourceforge.net/projects/uibm-de
In other pudeblications, please use:
Copyright © Apr. 2007
Uwe Grueters, Roland Dahlem, Jochen Senkbeil, Markus Woetzel
Some rights reserved
See http://sourceforge.net/projects/uibm-de
|
Navigator
|
You are here: Home
Local Navigator: Λ Top -> Next
Whole-Page Navigator: Home -> Background -> UIBM Development -> Virtual Experiments -> Outlook
|
Copyright © Sep. 2009 Dr. U. Grueters
|
|
|
Printable Version
|